Over the past decade, scientists discovered how bacteria defend themselves against viruses using CRISPR, a system that has revolutionized gene editing. But what if bacteria and viruses have been repurposing their own genetic tools for entirely different functions? A new study published in Nature by Columbia postdocs Tanner Wiegand, Egill Richard and Chance Meers uncovers a surprising twist in this evolutionary story: ancient genes, once used by viruses and mobile DNA elements called transposons, have evolved into RNA-guided transcriptional regulators – natural gene regulators that operate much like the immune system CRISPR, but without cutting DNA.
Bacteria are constantly in an evolutionary arms race with viruses. The mobile DNA pieces transposons have played a major role in bacterial evolution by jumping between locations in the genome and spreading genes. One of these genes, TnpB, was long thought to function only as a simple DNA-cutting enzyme, helping transposons move around. However, scientists have now discovered that some TnpB-derived proteins have lost their cutting ability and instead evolved into RNA-guided transcription factors – proteins that turn genes on or off by blocking access to DNA.
Wiegand and colleagues named this new class of proteins TldRs (TnpB-like nuclease-dead repressors). Unlike CRISPR proteins, which evolved from transposon-related genes to protect bacteria from viruses, TldRs appear to have been repurposed independently for gene regulation. These proteins rely on small RNA molecules to guide them to specific sequences in bacterial genomes, where they bind and shut down target genes by preventing transcription.
One of the most fascinating discoveries in the study is how viruses have co-opted TldRs to manipulate their bacterial hosts. The researchers found that some bacteriophages – viruses that infect bacteria – carry TldR genes along with a bacterial gene called fliC, which encodes flagellin, the protein that makes up the bacterial flagellum (a whip-like structure used for movement).
Normally, bacteria express their own version of fliC to build flagella, which are crucial for swimming and sensing the environment. However, in bacteria infected with certain bacteriophages, the viral version of fliC (fliCᴾ) replaces the host’s version. The study shows that TldRs, guided by their RNA molecules, specifically silence the bacterial fliC gene while allowing the viral version to be expressed. This means that the bacterium’s flagella are partially “rewired” to contain viral proteins instead of its own (Figure 1).
Why would a virus want to do this? In their study, Wiegand et al. propose a few possibilities:
- Evading the immune system – Many bacterial flagellins are recognized by immune cells, and replacing them with a different version could help bacteria (and their resident viruses) go unnoticed.
- Avoiding other viral infections – Some bacteriophages recognize flagella as entry points to infect bacteria. By altering flagellar proteins, the virus controlling the bacterium might block access to competing viruses.
- Affecting bacterial motility – Flagella are primarily involved in motility. Changing their structure could alter how bacteria swim in various environments.
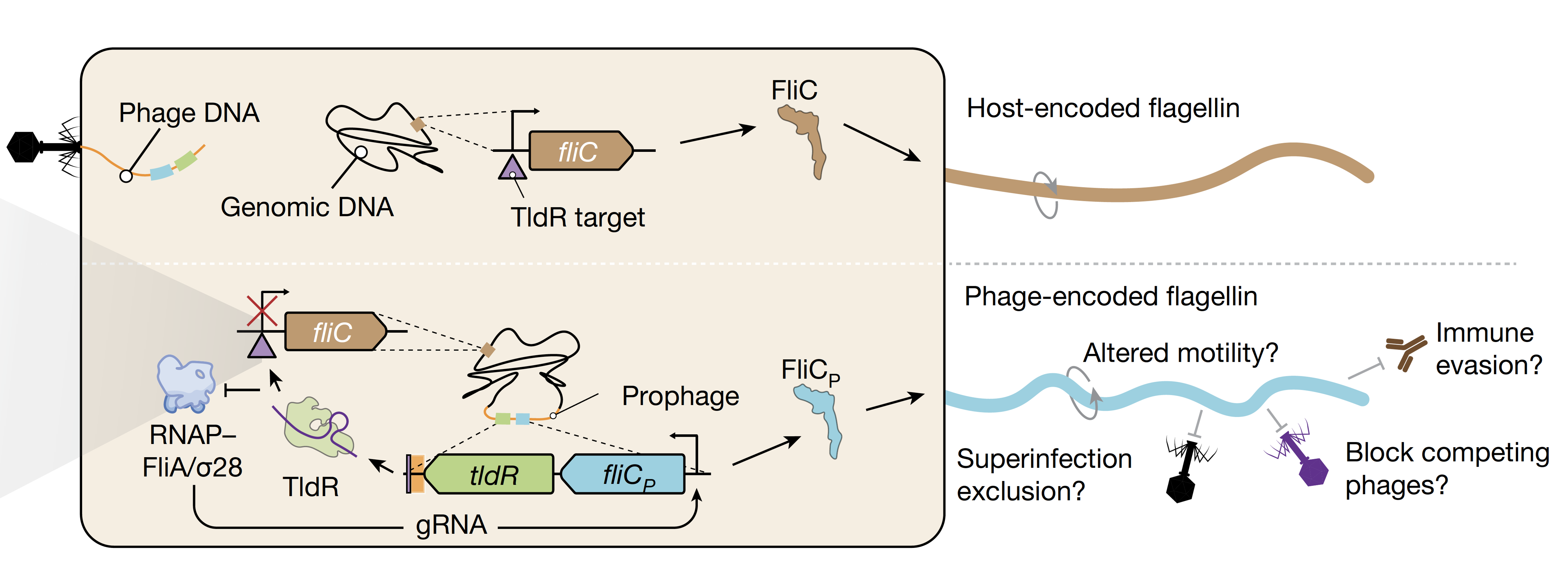
Figure 1: Schematic representation of the repression of the bacterial fliC gene by the RNA-guided viral-encoded TldR and consequent expression of the viral version of the flagellum gene, fliCp. Figure from the original paper.
One of the most exciting aspects of this discovery of this new class of gene regulators is their potential for biotechnology tools development. CRISPR-based gene editing relies on programmable RNA-guided proteins, and TldRs appear to work on the same principle – but without the need to cut DNA. Instead, they act as natural gene “dimmer switches”, fine-tuning expression levels in a targeted way. Because TldRs are much smaller than CRISPR-Cas proteins, they could be useful for future genetic engineering applications where space is limited, such as in gene therapy or synthetic biology. Unlike traditional CRISPR editing, which involves cutting DNA (which can lead to unwanted mutations), TldRs could provide a more precise way to repress or regulate genes without permanent genome changes.
This study is a striking reminder of evolution’s endless innovations and how nature continuously repurposes molecular tools over evolutionary time. The same transposon-derived genes that gave rise to CRISPR have now been independently adapted to regulate gene expression in bacteria. It also underscores how much we still have to learn about the diversity of RNA-guided systems beyond CRISPR, which could lead to new technologies inspired by nature’s own innovations.
With further research, TldRs might become a new class of programmable genetic tools, opening new doors in medicine, synthetic biology, and biotechnology. As we continue to explore these hidden layers of microbial evolution, who knows what other surprises nature has in store?
Reviewed by: Temistocles Molinar, Saheli Chowdhury